熱線:021-56056830,66110819
手機:13564362870
熱線:021-56056830,66110819
手機:13564362870
結果和討論
實驗性OA研究的主要特征
大多數調查僅限于一個個體發育階段(75%)、一代(96%)、一個因素(72%)、一個季節(96%)、一年(99%),是實驗室基于(73%)并以恒定強度(89%)應用治療(圖1)。例如,通過檢查對幾個個體發育階段的多因素影響同時對這些性狀中的兩個(或更多)進行升級的研究非常罕見(見表S1),即327個中的13個(或4%)。
由眾多單因素和單目標實驗產生的結果無疑在OResearch的“少年”階段很重要,這塑造了我們對給定pH變化對給定生物或給定亞生物過程的潛在影響的感覺。然而,就地而言,凈影響可能會有所不同。例如,壓力源在個體發育階段的影響可能是累加的、協同的或拮抗的(選擇性死亡率導致較舊階段的敏感性降低),并且僅知道pH值對一個階段的影響給出了不完整的圖景。此外,在一年中,多年生物種通常會經歷不同的生理階段,預計給定的pH值水平在不同季節會產生非常不同的影響。此外,原位pH值水平以及許多其他環境變量(Miller-Neilan和Rose 2014)在時間和空間上都是可變的(見下文)。浮游生物或多或少可以被認為是特定水體中的“居民”,除非它們垂直遷移,否則主要會隨著時間的推移經歷pH值波動。相比之下,當水運動(水流、向上或向下流)通過空間傳輸這種異質性時,定居的底棲生物會經歷時間波動和pH分布的空間異質性。
近年來,人們已經認識到其中的許多缺點。因此,OA越來越多地與其他環境因素相結合(例如Connell和Russell 2010;Connell等人,2013年;Hiebenthal等人,2013年;Duarte等人,2014年),OA效應在幾個個體發育階段被追蹤(例如Forsgren等人,2014年)。2013年;Kurihara等人,2013年),因季節而異(例如Miller等人2012年;McCoy 2013年)或跟隨了幾代人(例如Low-De′carie等人2011年;Lohbeck等人2013年)。然而,與自然棲息地類似的頻率和振幅的pH值或pCO2波動如何調節相對于恒定條件的OA影響,實際上仍未得到探索。一個可能的原因是,大多數棲息地不存在對這些原位波動的長期、高分辨率監測。
碳酸鹽化學的原位波動
在下文中,我們將展示最近發表的關于不同尺度的空間和時間pH波動的證據。我們將僅簡要提及這些波動的驅動因素,因為Waldbusser和Salisbury(2014年)最近對這些因素進行了出色的審查。
總的來說,我們發現了54篇出版物,其中至少有一個碳酸鹽化學描述詞(pCO2、pH、總堿度、溶解無機碳(DIC)、[CO2](aq)、[HCO3–]、[CO3 2?])進行了原位監測。我們注意到底棲棲息地之間的研究工作存在很大差異。迄今為止,熱帶珊瑚礁的24項研究(占研究總數的49%)受到了大部分研究關注(表1)。九項研究(占總數的16.5%)代表了對潮間帶巖池波動的研究。自1950年代以來,巖石池就被認為是pCO2和O2波動極大的環境(Daniel和Boyden 1975;Truchot和Duhamel-Jouve 1980),并且特別引起了生理學家的注意。海草草甸也只有9項研究(16.5%)代表,考慮到它們的生態重要性,這是一個很小的數字。大多數這些研究是在地中海的Posidonia Oceanica草地上進行的(Frankignoulle和Diste`che 1984;Frankignoulle 1988;Frankignoulle和Bouquegneau 1990;Hendriks等人,2014)。盡管Zosteraceae廣泛分布于溫帶地區(Buapet等人,2013年),但僅發現了一項關于Zosteraceae的研究,并且只有兩項研究側重于Thalassia spp的熱帶海草草甸。(Yates等人,2007年;Semesi等人,2009年)。海藻棲息地(不包括海帶,占總數的11%)以及海帶森林(占總數的7%)相對于其生態相關性的研究嚴重不足。此外,在此背景下的所有海帶研究都是在Macrocystis pyrifera棲息地進行的(Delille等人,2000年,2009年;Hofmann等人,2011年;Frieder等人,2012年)。我們沒有發現任何關于溫帶或熱帶腹足動物或雙殼類珊瑚礁(或由熱帶造礁珊瑚以外的生物形成的任何珊瑚礁)的研究,例如由牡蠣或貽貝形成的珊瑚礁。確認底棲生態學的總體趨勢,我們發現研究工作強烈傾向于熱帶珊瑚礁。
海水pH值可能會在局部范圍內(公里到亞毫米)發生生物學改變,尤其是在以海草、大型藻類、貽貝床、珊瑚或其他工程底棲生物為主的沿海系統中(Duarte等人,2013年回顧)。主要機制是微藻和大型藻類在光合作用過程中吸收CO2(或HCO3–),所有生物在呼吸過程中釋放CO2,鈣化劑在殼形成過程中降低總堿度和釋放CO2(例如Hurd等人.2009)。鈣化過程中釋放CO2的機制如下:HCO3–解離為Ht和CO3 2?,而后者作為骨架CaCO3不斷沉淀,Ht再次與HCO3結合-并形成H2CO3,隨后分解為H2O和CO2(Hurd等人,2009年)。因此,自養生物、異養生物和鈣化生物的光合作用、呼吸作用和鈣化活動之間建立了微妙的反饋回路平衡。這些過程受生物體的生理狀態和能量可用性以及各種非生物因素的影響,例如溫度、光和養分的可用性以及水的運動。此外,水運動將不同預處理的水體輸送到特定的棲息地,例如通過垂直對流、潮汐、洋流和上升或下降。所有這些因素反過來會隨著一天中的時間、天氣條件、季節甚至年際而波動,最終導致流入或流出生物體的凈碳流量的時間或空間反轉,從而導致海水pH值的生物波動。預計這些波動的幅度與生物量與水的體積比呈正相關,與水交換率呈負相關,而頻率和幅度在空間和時間尺度上都可能發生很大變化。pH值的生物調節可能足以完全掩蓋沿海棲息地海洋酸化的全球趨勢(Anthony等人,2013年;Duarte等人,2013年)。
表1.碳酸鹽系統自然波動的原位研究數量數據基于54篇出版物(見表S2)。括號中的數字是百分比
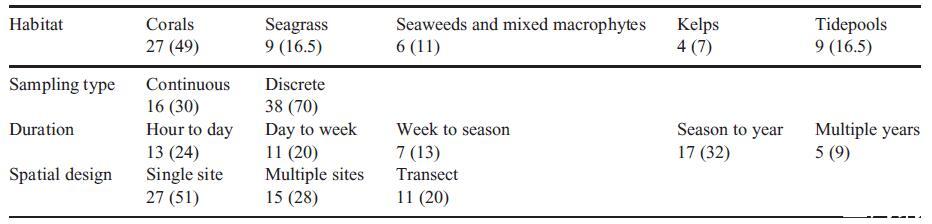
大尺度時空波動:年際變化、年代際循環、氣候變化
碳酸鹽化學的多年變化受到的監測最少。在底棲棲息地進行的長期系泊記錄很少,例如珊瑚礁(Drupp等人,2011年,2013年)。這些緩慢的波動可能會受到周期性氣候事件的影響,例如北大西洋濤動的變化或厄爾尼諾-拉尼娜序列的變化。pCO2的多年定向變化,如在氣候變化過程中發生的(Stocker等人,2013年),不是嚴格意義上的波動,也不是本次調查的主題。應該提到的是,這種長期變化在小空間尺度(例如,巖石池中-0.039到-0.054 pH單位年?1)可能比公海(-0.0019 pH單位年-1)發生得更快)(伍頓等人,2008年)。
中尺度時空波動:季節
季節性,特別是由于光和溫度的變化,對海水化學有很大影響(Frankignoulle和Diste`che 1984;Delille等人2009;Shaw等人2012)。光周期和輻照度的變化影響生態系統的自養與異養平衡,溫度影響代謝活動,這兩個因素隨著緯度的升高而增加。據報道,亞南極地區(凱爾蓋朗群島、法屬南部和南極土地)的海帶森林(Macrosystis pyrifera)的pCO2季節性變化幅度為,500 matm,冬季的晝夜變化約為夏季的一半(Delille等人,2000年,2009年)。在溫帶地區,Frankignoulle和Diste`che(1984年)報告了地中海(法國科西嘉島)海草草甸(Posidonia Oceanica)中pCO2的季節性變化為500平方米。在夏威夷珊瑚礁(瓦胡島)中,Drupp等人。(2011)發現pCO2的季節性變化幅度為61 matm,季節之間的晝夜幅度沒有明顯變化。在百慕大附近的珊瑚礁中,Bates和Leone(2001)發現季節性變化為100 matm。
季節性通常還表現為氣象事件強度的變化,例如雨和風情(Manzello 2010;Saderne等,2013)。雨季降雨和河流徑流增加會稀釋海水,從而降低鹽度,從而降低海水的總堿度和緩沖能力。在這種情況下,我們預計生物源pH值變化的幅度將最為顯著——如果生物體的活動不會因這種潛在的滲透壓力而降低(例如,Saderne等人,2013年)。在熱帶潮濕季節的巴拿馬珊瑚礁中觀察到了這種影響(Manzello 2010)。緩沖能力降低也可能解釋了在半咸水環境(例如Elkhorn河口(Hofmann等人,2011)或波羅的海西部(Saderne等人,2013年)中記錄的特別重要的晝夜pH值變化)。然而,在另一個方向上,河流徑流引入的溶解有機質負荷可以通過增加堿度來增加緩沖能力(參見Kulin′ski等人,2014年的解釋)。除了對緩沖能力的這些影響之外,河流徑流還會影響海水溫度、注入營養物質(磷酸鹽、硝酸鹽和硅酸鹽)并增加濁度。總之,這些因素的結合會在季節性或風暴事件后影響生物的生物學和海水的化學性質。例如,在一些珊瑚礁中發現了這種情況,那里的碳酸鹽化學在大風暴之后發生了變化(Massaro等人,2012年;Drupp等人,2013年)。
由氣象學驅動的瞬時上升流是影響沿海棲息地碳酸鹽化學及其動力學的另一個重要現象。在上升流期間,溫躍層的淺灘給淺海沿岸棲息地帶來了更冷、通常更多、pH值和pO2較低、pCO2較高的咸水團。這些水團與大型植物光合作用的相互作用可以在碳酸鹽化學的生物成因變化中產生極端變化。Saderne等人證明了這一點。(2013)在季節性上升流到波羅的海西部的大型藻類棲息地期間。在此事件期間,大量海水pCO2從大氣平衡(426 matm)增加到1600 matm,同時鹽度增加0.2個單位,溫度下降0.38C。大量水非生物參數(pCO2、鹽度和溫度)變化的結合導致晝夜節律pCO2振幅增加到600 matm,并導致0.35個單位的pH值變化的反直覺下降。這些影響不是由于光合作用增強(減少了20%),而是由于CO2、HCO3-2和CO3-2之間的平衡發生了變化。
小尺度時空波動:晝夜循環和日間變化
在小型沿海棲息地或浮游生物斑塊的晝夜循環中觀察到最顯著的pH值或pCO2波動模式。在這個小的時空尺度上,振幅很大。這主要是由于24小時內輻照的振蕩導致白天通過光養生物高凈CO2吸收,這會增加pH值,而所有生物在夜間釋放CO2會降低pH值(Koch等人,2013年)。這些晝夜波動的幅度可能因季節而異。例如,在海藻林中,據報道,pCO2在晝夜循環中的變化在冬季為160 matm,在夏季為340 matm(Delille等人,2009年)。同樣,溫帶墨角藻中pCO2的晝夜波動。站在夏末的振幅在240到400 matm之間,秋季可能達到2000 matm(Saderne等人,2013年)。由于天氣條件和浮游生物遮蔽,UW輻射的更多短期和零星波動預計會導致大型植物林分中pH值波動的頻率和幅度發生相應變化。除了光之外,由于潮汐周期或風事件,海流的速度和方向可能會在一天內發生變化。低流速會降低水體的交換率和稀釋度,這應該允許pH值變化幅度更大,而高流速會導致幅度降低。死水的晝夜波動最為強烈,例如,它發生在潮汐池中。在這里,經常發現pCO2的日變化超過1000 matm day?1(Truchot和Duhamel-Jouve 1980;Morris和Taylor 1983;Nguyen和Byrne 2014)。電流速度隨著到襯底的距離的減小而減小。Shashar和他的同事(1996)從米的尺度放大到基質上方的微米,區分了三個邊界層層:底棲邊界層、動量邊界層和擴散邊界層。底棲邊界層是由水與底層浮雕、藻類和海藻冠層或礁石摩擦產生的,尺度從米到厘米(Hurd 2000)。它導致水在底棲生境中的停留時間增加,從而導致碳酸鹽化學變化幅度的增加。由基質、植物或無柄底棲動物的地形,以及它們的泵、纖毛或觸手活動產生厘米到毫米尺度的動量邊界層,產生非常局部的小尺度電流(Shashar et al.1996)。例如,康沃爾等人。(2013)描述了從覆蓋有關節珊瑚藻的鵝卵石組合上方10厘米到距藻類表面幾毫米以內的pH值日夜幅度越來越寬。在1.5 cm s?1的中等體積水流速下,晝夜條件之間的pH值差異在距離藻類表面68毫米處為0.05,在距離藻類表面6毫米處幾乎為0.3。擴散邊界層是由于水的粘度在代謝活躍組織表面形成的分子的微尺度濃度梯度。在一項未發表的研究中,我們研究了褐藻Fucus vesiculosus葉狀體上方擴散邊界層中的pH梯度。我們發現在死水條件下,Fucus葉片表面的暗光pH值波動在幾分鐘內超過1.0個單位。在增加體積水速度(0、0.5和1.5 cm s?1)下,Fucus vesiculosus thallus的邊界層厚度從700毫米減少到200毫米(速度0.5和1.5厘米s?1之間沒有差異)和振幅邊界層內pH值的暗-光波動減小(圖2)。在最接近基質的地方,在停滯條件下pH值波動1.5個單位,在中等水流下波動0.5個單位(圖2)。因此,生活在由初級生產者主導的這種微型或納米棲息地中的生物將經歷強烈而快速的pH變化,這可能會在幾分鐘內H+離子濃度增加或減少30倍(圖2)。例如,根據它們的體型,Fucus thallus上的表生生物將在邊界層的不同地層中經歷不同的波動狀態。苔蘚蟲或幼年藤壺(500毫米高)會受到微弱的pH值波動(0.2單位),并且僅在停滯條件下。許多無脊椎動物(通常高200-250毫米)的新鮮幼蟲在停滯條件下會經歷強烈的波動(1個單位),而在流動條件下則沒有波動。硅藻(1-50毫米)必須承受強烈的波動(停滯時為1.3個單位,流動條件下為0.3個單位)。最后,細菌(0.5-5毫米高)在流動條件下在停滯和中等(0.4個單位)波動下會受到極端波動(1.5個單位)。應該注意的是,這些值是在黑暗和光明之間切換后5分鐘測量的。這意味著在停滯條件下,而不是在流動條件下,邊界層的厚度可能會隨著時間的推移而增加。此外,溫度會影響水的粘度和氣體的溶解度,從而影響海水的化學性質。例如,在擴散邊界層的情況下,這意味著較高的溫度會導致較低的粘度,這會導致除停滯之外的所有流態中的邊界厚度減小。此外,擴散速率和代謝速率增加,導致在較窄的邊界層內總體上出現更陡峭的pH梯度。然而,這仍然需要測試。
到墨角表面的距離(μm)

圖2.棕色巨藻Fucus vesiculosus葉狀體上具有擴散邊界層的pH梯度。在測量前5分鐘內,允許在明暗條件下以各種體積速度建立擴散邊界層。葉綠體表面位于距離=0處,即位于圖的右邊界處。垂直線描繪了大型藻類葉狀體表面(“0厘米”)上方的地層的外部界限,不同的表生生物通常生活在其中。
所描述的pH值的強烈波動可能代表壓力階段和壓力釋放階段的交替。直接環境的這種生物不穩定性可能對生理性能有益或有害。它是否有潛力局部調節全球pCO2(OA)穩步上升所施加的假定壓力,目前尚不清楚。
空間尺度與效果大小(幅度)之間的關系
鑒于較大水體的慣性越來越大,我們預計生物波動的幅度與空間尺度成反比。珊瑚礁等沿海棲息地的生物碳酸鹽化學波動幅度確實與水柱大小和水交換率成反比(Bates和Leone 2001)。例如,Ohde和van Woesik(1999年)在沖繩島(日本)附近環礁的橫斷面研究中發現,環礁中心最淺的區域(礁坪)的最大晝夜振幅為927米。潮汐池的水柱高度和交換率降低,是生物活動產生特別強烈的碳酸鹽化學變化的棲息地,波動幅度隨著海岸高度的增加而增加(Daniel和Boyden 1975)。迄今為止,生物源pH值波動的最大振幅記錄在非常小的尺度上(即在亞毫米尺度的底棲邊界層中)(Spilling等人2010;Hurd和Pilditch 2011;以及上面給出的例子)。pH值波動的空間尺度、頻率和幅度之間的反比關系如圖3所示。由于較小的水體具有較小的慣性,因此尺度和變化率不是獨立的。事實上,pH變化率與所考慮的空間尺度呈負相關(圖4)。最慢的變化發生在公海,在沿海棲息地觀察到快速變化,最劇烈的pH值波動來自藻類表面邊界層中亞厘米甚至亞毫米的“納米棲息地”(Spilling等人,2010年;Cornwall等人)2013年)。在開闊的海洋中,浮游生物斑塊(Schulz和Riebesell 2013)發生幅度小得多的晝夜和季節性波動,在這種情況下,可以將其視為漂浮的微生境。
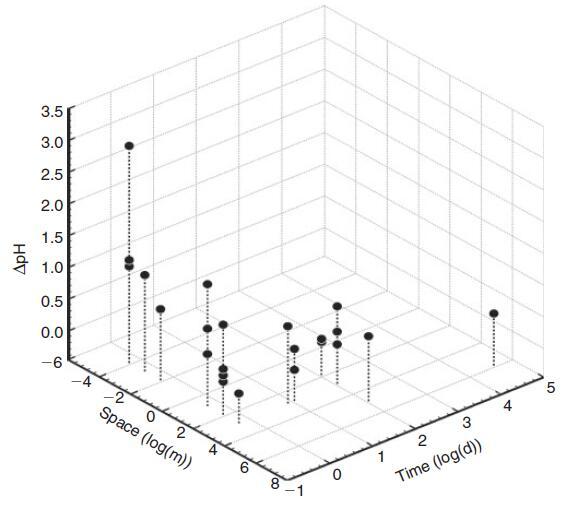
圖3.pH波動的典型振幅('delta pH')和頻率('Time')與所考慮系統的估計大小('Space')之間的關系。數據是從報告原位波動的選定論文中提取的(Morris和Taylor 1983;Middelboe和Hansen 2007;Semesi等,2009;Spilling等,2010;Thomsen等,2010;Hofmann等,2011;Hurd和Pilditch);Frieder et al.2012;Gray et al.2012;Price et al.2012;Cornwall et al.2013;Johnson et al.2013;Melzner et al.2013;Saderne et al.2013;Saderne and Wahl Stock et al.2013;al.2013;Comeau et al.2014;Hendriks et al.2014)以及本文中作者自己的測量結果。需要注意的是,這些文章中的幅度、時間和空間尺度是根據這些文章中的圖形和文本信息估算的,僅代表數量級。
波動的生物效應
我們對pH值或pCO2波動的生物學相關性的普遍無知源于前面提到的方法學研究局限性。我們在實驗室進行的大部分調查都使用恒定的處理水平。在現場實驗中,可以容忍波動,但很少評估它們相對于非波動條件的重要性。在這一點上,重要的是要注意恒定和波動條件不是兩個離散條件。相反,“恒定性”代表連續體上的一個點,其中變量波動的頻率相對于生物體的生物節律較長,或幅度接近于零。
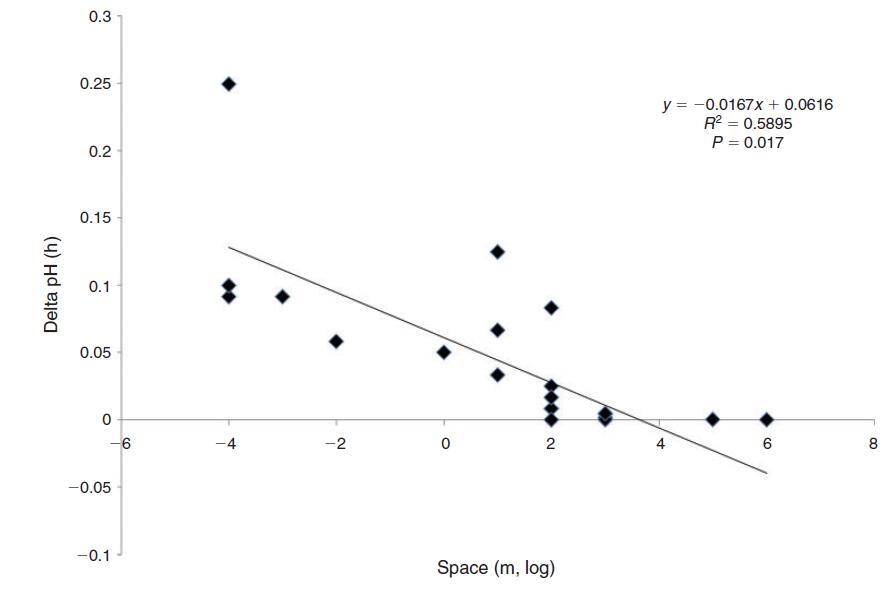
圖4.不同空間尺度下pH值的變化率。考慮的尺度越小,單位時間pH變化的斜率越陡,即pH變化越快越強。線性回歸結果顯示在右上角。數據來源如圖2所示。
波動的可能影響有很多。顯然,在波動的情況下,與環境變量(相對于目標物種的要求)的最佳值的距離不斷變化。因此,在這種波動期間,物種的相對性能將增加或減少。生理反應可能會顯示一些滯后,這反映了生物體的生理緩沖(例如內部pH控制)。當波動的變量周期性地超過生理閾值時,壓力階段可能與壓力放松階段交替出現。波動的制度可能會選擇能夠應對各種環境變量的“生理通才”。盡管許多生物體在對波動的環境變量的反應中表現出顯著的表型可塑性(Hadfield和Strathmann 1996),但這種可塑性可能需要付出一些代價(Waldbusser和Salisbury 2014)。當環境參數變化快時,可塑性可能太慢,當波動具有高幅度和高頻率時可能會發生這種情況。當生物體無法適應快速的瞬態變化時,它們的耐受范圍應該很寬,否則它們可能會關閉敏感過程。當預期的波動包含在變暖處理中時,對變暖的反應更強烈,更消極(例如Vasseur等人,2014年)。一些這種增加的影響可能歸因于Jensen不等式(圖5)預測當響應曲線是曲線時,變量的對稱波動通常會產生生物體的不對稱響應。波動制度的另一個壓力方面是極端事件更有可能發生。耐受閾值的瞬時越界,隨后是松弛期,不僅可能有選擇地有利于高表型可塑性(見上文),而且還可能選擇更強大的基因型(Pansch等人,2014年)。
只有少數研究調查了pH(實際或未來)在相同平均pH(或pCO2)的恒定與波動pH方案中對生物體的影響是否不同。鈣化大型藻類似乎在低pCO2水平和升高的pCO2水平下都會受到pH值波動的影響(Cornwall等人,2013年)。然而,厚邊界層可能允許光合鈣化劑在其葉狀體表面創造有益的納米棲息地,即使大量水被酸化到臨界水平,也允許鈣化(Cornwall等人,2014年)。這些作者表明,導致較薄邊界層的強烈水湍流可能會削弱這種保護功能。低pH值脈沖(模擬上升流)會降低蝸牛的性能(Kim等人,2013年),但這種影響并未與具有相同平均pH值的恒定方案進行比較。波動的pCO2制度有利于珊瑚新兵的生長,可能是因為當pCO2高時,它們更容易隔離DIC(Dufault等人,2012年)。pH值波動可能會緩沖海洋酸化對珊瑚鈣化的負面影響(Comeau等人,2014年)。當環境波動時,兩種貽貝物種的幼蟲受海洋酸化的影響較小(Frieder等人,2014年)。同樣,Schneider、Sawall、Saderne、Hiebenthal、Müller和Wahl(未公開數據)發現pH值的生物波動減輕了海洋酸化對貽貝鈣化的影響,因為貽貝將其鈣化活動從酸化壓力(即白天)。
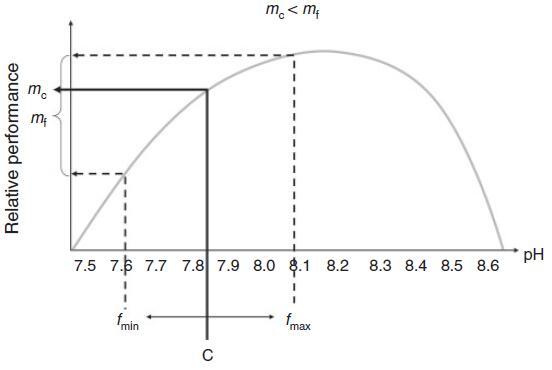
圖5.Jensen不等式說明當環境變量和生物體性能之間的關系是非線性時,相同平均值的恒定(c)和波動(f)狀態可能會產生不同的響應。因此,圍繞驅動器平均值(c)的對稱波動(在fmin和fmax之間)(本例中為pH值)可能會導致對恒定pH值的平均響應(mc)周圍的不對稱響應分布(mf),因此,降低目標物種的性能(mf,mc)。
已經描述了其他環境因素的一些波動效應。相對于相同平均值的恒定條件,脈沖低鹽度對植物Halophila的壓力更大(Griffin和Durako 2012),光脈沖減輕浮游植物的低光照壓力(Helbling等人,2013),以及溫暖的平均壓力珊瑚周圍的溫度波動比恒溫狀態更嚴重(Putnam和Edmunds 2011),但恒溫狀態稍涼!)。在一項理論研究中,Vasseur及其同事(2014年)提出,相對于恒定的溫度狀況,波動有利于生活在較冷分布邊緣的物種,但不利于生活在較暖分布邊緣的物種,并且全球變暖的影響可能比想象的更劇烈一種將波動融入畫面中。這些假設的經驗證據仍然缺乏。
 相關新聞
相關新聞